 |
|
 |
| Paleoanthropology | ||
|
|
||
| (revised 20 February 2007) |
|
|
|
| Paleoanthropology | ||
|
|
||
| (revised 20 February 2007) |
Contents
|
Introduction: The Hominid Family top The terminology of our immediate biological family is currently in flux; for an overview, see a current hominoid taxonomy. The term "hominin" refers to any genus in the human tribe (Hominini), of which Homo sapiens (modern man) is the only living specimen. We don't have to go too far back into the past, however, to find relatives (cf. "We Were Not Alone," SciAm Jan 2000). Discounting abominable snowmen, yeti, bigfoot, and other merely rumored possible members of our family, we know that only 28,000 years ago Neanderthals still thrived in Europe. More surprisingly, recent evidence (see below) suggests that a member of even longer standing, Homo erectus, who first appears in the fossil record nearly two million years ago, may have continued to inhabit the island of Java as recently as ten thousand years ago, or into historical times. Looking further back, the various members of the genus Homo are the decendants of the australopithecines, who around three million years ago were represented by numerous species. These clearly belonged to the great apes: their brains were about the size of chimpanzees, we have found no record of their tool manufacture, and they were probably sexually promiscuous in the way chimpanzees are. There were also about the same size as chimps: Australopithecus afarensis weighed around 25 to 50 kilograms (60 to 120 lbs.) as an adult. The main difference between the two species is that the australopithecines walked upright - the first record of their footprints was discovered at Laetoli in East Africa and has been dated at 3.6 million years. Its recently discovered ancestor, Ardipithecus ramidus, retained more adaptations for climbing, as seen from the shape of the limbs. Its dates are very close to the split between the chimpanzees and the hominids; a find still closer is the recently discovered and still debated Orrorin tugenensis (below). For a recent overview of all the different members of our family, see a discussion of hominoid taxonomy, Donald Johanson's Hominid Family Tree, and a simplified tree situating the new A. garhi (cf. below). While several of these extinct and living species are acknowledged to be our relations, the precise details of descent remain contentious. Note, for instance, that the discoverers of Orrorin (below) argue that Orrorin is our ancestor, while ardepithecus is the ancestor of Pan (the chimpanzees) and the Australopithecines went extinct (details in Orrorin tugenensis), while the discoverers of . Based on molecular evidence, the current consensus is that our closest living relatives, the two species of Pan (the chimpanzee and the bonobo), separated from the hominid lineage between five and six million years ago. While our line diverged into several species of australopithecines and then several species of homo, Pan separated into two species starting around a million years ago. Hominids, gorillas, chimpanzees, and bonobos - the four African species of Great Apes - have common ancestors somewhere between eight and ten million years ago (BBC report, external). For our living relatives, see the recent detailed tree showing humankind's relationship to the African apes. Looking even further back, see current speculations on the origins of the anthropoids. For primates in general, see Primate Info Net (external). For detailed articles on the various members of the hominid
family, see the external site Origins of
Humankind. See also the quick
overview at Talk Origins. Rapid evolution of nervous system genes The study of hominids have given a special place to our own ancestors, creating what appears to be a distorting factor in paleoanthropological interpretations. Since there is a relatively low number of relevant fossil finds, new finds often create an opportunity for reinterpreting the existing data, and this reinterpretation appears at times to favor placing one's own remains at the root of the human tree, rather than in the line of descent of the chimpanzees, our closest living relatives. Yet two recent finds from the period close to our split from the chimpanzees remain interesting whether they are our own ancestors or not. Orrorin tugenensis, the Lukeino hominid top The Kenya Palaeontology Expedition (KPE) report in December 2000 the discovery of what is almost certainly a new species of hominid at Kapsomin in the Tugen hills in Kenya's Baringo district; see the BBC news story. The remains were found in volcanic sediments dated to between 5.6 and 6.2 million years old. They include a left femur, pieces of jaw with teeth, isolated upper and lower teeth, arm bones, and a finger bone; the excavations are ongoing. Preliminary analyses suggest the hominid, the size of a chimpanzee, was an agile climber and that it walked on two legs when on the ground. The tentative date of six million years indicate a date very close to the common ancestor of humans and chimpanzees, although this date may now need to be pushed back. The excavating team includes Martin Pickford from the KPE and Brigitte Senut from the Museum of Natural History in Paris. The find was published in January 2001, see details and discussion (local). Ardipithecus ramidus top A related find from close to the same period (5.2 to 5.8 million
years ago) is reported in Nature in July 2001; see report in
Science 293. 5528 (13 Jul 2001): 187-189 (full
text, external). It was discovered in 1997 by an
Ethiopian-American team led by Yohannes Haile-Selassie and Tim White
of the University of California, Berkeley. The specimen has been
named Ardipithecus ramidus kaddaba, a species of the
Ardipithecus ramidus first discovered in 1994. While Senut and
Pickford (2001) propose that Ardipithecus is the ancestor of Pan, the
chimpanzees (see details), Haile-Selassie
and White (2001) argue it is more likely to be a human ancestor. They
add that ongoing excavations may locate further remains to help
determine the issue.
Australopithecines top
Dean Falk and coworkers have recently examined australopithecine endocasts. In one of the four australopithecine species that were studied, certain parts of the cerebral cortex appeared advanced compared to the brains of living great apes. "Endocasts from Australopithecus africanus reveal that its brain size was, on average, within the range of living chimpanzees. The underneath surfaces of its frontal lobes and the anterior tips of its temporal lobes, however, appear to have evolved to a point that was intermediary between the forms seen in great apes and humans." This may be indicative that we descend from the gracile australopithecines. Falk, D., Redmond, J.C., Jr., Guyer, J., Conroy, G.C., Recheis, W., Weber, G.W. and H. Seidler. Early hominid brain evolution: A new look at old endocasts, Journal of Human Evolution, Spring 2000. Author's presentation. Bipedalism top The most significant adaptation that distinguishes our first hominid ancestors from the chimpanzees is upright walking, or bipedalism. The adaptation strictly speaking predates the australopithecines. Various theories have attempted to explain the trait, the simplest being that upright walking freed the hands for carrying objects. Still, it is not clear why this capacity would matter enough to drive such a major adaptation. In Human
evolution: the water theory (1998, external), Elaine Morgan
discusses an alternative theory, arguing that bipedalism may have
arisen during an aquatic stage. The hypothesis is intriguing but hard
to test. For a discussion of the significance of running for the
transition from the Australopithecines to Homo erectus, see "Endurance
running and the evolution of Homo" below. Bipedalism had the effect of opening up an evolutionary possibility space for future adaptations such as tool use and possibly enlarged brains (see below, What drove the transition to Homo?) and speech. Robert Provine, in his book Laughter (Little/Brown, spring 2000), argues that bipedalism was a necessary condition for language: In quadrupeds, there is a one-to-one correlation between breathing pattern and stride because the lungs must be fully inflated to add rigidity to the thoracic complex (sternum, ribs and associated musculature) that absorbs forelimb impacts during running. Without such synchronization, the thorax is weak and unable to absorb the impact. When primates stood and walked on two legs, the thorax was freed of its support function during locomotion, breaking the link between breathing patterns and stride. This flexibility enabled humans to regulate breathing and ultimately, speak" (press release). Australopithecine predation Raymond Dart's discoveries of australopithecine fossils in
South Africa's limestone caves, starting in the 1920s, led him at first
to propose a theory of a "killer ape". Further reseach showed that the
bones were evidence of predation, possibly by felines. More
recently, Lee Berger, a paleo-anthropologist at
Johannesburg's University of Witwatersrand, has argued large birds
preyed on these apes. Eagles have been documented to kill monkeys by
piercing the skull behind the eyes, and Berger found hitherto unnoticed
marks in Dart's spectacularly well-preserved Tuang baby (see 12 Dec
2006 news report). Australopithecus garhi top A new species of australopithecine was discovered in Ethiopia in 1999, dated to about 2.5 million years and in certain ways anatomically intermediate between A. afarensis and Homo habilis. What makes the find particularly interesting is that the archaeologists also found suggestive evidence that this type of early hominid used tools and ate meat. Its placement in the family tree is not settled; see diagram. Berhane Asfaw, Tim White, Owen Lovejoy, Bruce Latimer, Scott Simpson, and Gen Suwa. Australopithecus garhi: A New Species of Early Hominid from Ethiopia. Science 284 (April 23, 1999): 629-635. Media reports (local copies): Fox, Maggie. Meat-Eating Missing Link Fossil Found In Africa. Reuters, April 22, 1999. Wilford, John Noble. Skull and Fossils Found in
Ethiopia Prompt Missing-Link Debate. New York Times, April
23, 1999. Australopithecine Diet top Little is known about the diets of hominids that predate the Homo genus, because these hominids did not leave archeological traces such as "kitchen middens" and stone tools. Consequently, researchers have made inferences concerning hominid diet on the basis of craniodental morphology, gross dental wear, and dental microwear. The current consensus is that the 3-million-year-old Australopithecus africanus hominid subsisted on fruits and leaves, similar to the modern chimpanzee. Early hominid diets are of some theoretical significance, since one current view is that the emergence of the more intelligent Homo genus depended on the consumption of high-quality animal foods that made possible biological changes resulting in the evolution of a larger brain. M. Sponheimer and J.A. Lee-Thorp (1999) now report a stable carbon isotope analysis of A. africanus fossils from Makapansgat Limeworks, South Africa. The authors sampled 4 of the 14 Australopithecus africanus individuals that have been unearthed at that location, and also analyzed the dental enamel of associated 3-million year old animals (65 individual animals from 19 mammalian taxa) in order to place A. africanus within a broader ecological context. The authors report their results demonstrate that A. africanus ate not only fruits and leaves, but they also ate large quantities of carbon-13 enriched foods such as grasses and sedges, or they ate animals that ate these plants, or both. The authors suggest their results indicate that early hominids such as A. africanus regularly exploited relatively open environments such as woodlands or grasslands for food, and that early hominids may have consumed high-quality animal foods before the development of stone toolsand the origin of the genus Homo. ( Science Week April 1999) M. Sponheimer and J.A. Lee-Thorp (1999). Isotopic evidence for the diet of an early hominid, Australopithecus africanus. Science 283 (15 Jan): 368. Contact: Matt Sponheimer, Rutgers University, New Brunswick
908-932-8789. Peter Ungar of the University of Arkansas analyzed molars from
early homo (2.5 mya) and Australopithecus afarensis, concluding that
homo had switched to a tougher diet, most likely meat.
Australopithecine teeth suggest a soft diet of fruits. New Scientist, 3 Sept 2003. News report (external). Dr Lucinda Backwell (Witwatersrand University, Johannesburg)
report in Proceedings of the National Academy of Sciences
(early 2001) that A. robustus likely used bone tools to dig for
termites; see news
report (external). What drove the transition from Australopithecus to Homo erectus? top The Australopithecines were a diverse set of species that thrived
for several million years with little or no change in brain volume.
What were the factors that led to the evolution of the much brainier
Homo erectus? Some researchers emphasize a possible transition
to hunting and meat-eating. Even though chimpanzees also hunt and
there is now evidence that Australopithecines ate meat (see previous
two sections), tool-based hunting may have created a novel set of
adaptive pressures. Another possibility currently being raised by Richard
Wrangham et al. is that late australopithecines began to cook
tubers, which are and were plentiful on the East African savannah,
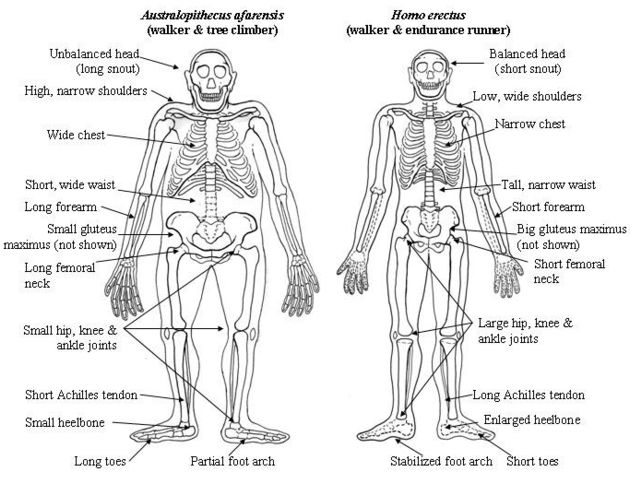
and that this created a new dynamic that led to larger brains; see Pennisi's presentation and the recent evidence of controlled fire from 790,000 years ago. O'Connell et al., in an article in the summer 1999 issue of the Journal of Human Evolution, muster evidence to support the tuber hypothesis. It is generally agreed that a 'grade shift' -- a conjunction of significant innovations -- in human evolution can be found in the fossil record of a little less than two million years ago. Here we see the first signs of hominins (human ancestors from the tribe Hominini) whose bodies, jaws and teeth begin to resemble those of modern humans, the type broadly labeled Homo erectus. O'Connell and colleagues challenge conventional wisdom by suggesting that the cascade of changes - involving anatomy, life history, culture and social structure - that resulted in the evolution of modern humans was set off not by hunting, but by systematic exploitation of subterranean food sources and recruitment of grandmothers to provide for their grandchildren. Larger brains have plausibly been connected with the evolution of a distinct human sexual psychology, favoring pair-bonding over promiscuity. Among both species of chimpanzees, females have exclusive charge of infants. Because of promiscuous sexual practices, paternity is generally not trackable. Australopithecines have similar brains and bodies and we have as yet no reason to think they didn't follow similar practices. A selection for larger brains -- for whatever reason -- would run up against the problem of a baby's larger head needing to pass through the bones surrounding the birth canal. The solution in place today is that human babies are born very prematurely compared to the offspring of our closest relatives. The still soft head of a new-born infant is deformed into a tube as it is squeezed through the birth canal. The price humans pay for this is a requirement of increased maternal care; for instance, for many months the baby is completely incapable of any form of locomotion, or even of clinging to the mother. In addition, larger brains would have favored extending the period in which infants are primarily devoted to learning new skills. These factors are likely to have created a significant benefit for children who received care not only from their mothers but also from their fathers and possibly their grandmothers (in the latter case creating a selective pressure for menopause). In the case of fathers, natural selection would have favored males who were able to invest in their own rather than in others' offspring, thus creating selective pressures some way for males to track paternity. Pair-bonding is the likely solution -- one that necessitated novel psychological adaptations, possibly along with physiological ones such as continued sexual receptivity in females. A large class of adaptations spring from this matrix of problems arising from larger brains. The dramatic increase in brain size from australopithecines to Homo erectus may represent the beginning of this process. In "Endurance running and the evolution of Homo"
(Nature 432, 345-352; 18 November 2004), Dennis M. Bramble and Daniel
E. Lieberman write, "The fossil evidence of these features suggests
that endurance running is a derived capability of the genus Homo,
originating about 2 million years ago, and may have been instrumental
in the evolution of the human body form." They argue that running
explains some of the anatomical changes that characterize the
transition from the Australopithecines to Homo erectus, as shown in the
drawings below.
Australopithecus afarensis compared to Homo erectus.
Where to Draw the Genus Boundaries top In a recent article, Wood and Collard propose a revised definition for the genus Homo based on criteria considered verifiable. They suggest that a fossil species should be included in Homo only if the following can be demonstrated:
The adoption of the above criteria, the authors conclude, would mean the genus Homo would have both phylogenetic and adaptive significance. "Researchers can then explore whether this adaptive shift in hominin evolution corresponds with changes in climate, analogous evolutionary changes in other large mammal groups, particular innovations in the hominin cultural record, substantial expansions in geographic range, or changes in ecological tolerance as reflected in reconstructions of hominin habitats." B. Wood and M. Collard: The human genus. Science 284 (2 Apr 99): 65. Contact: Bernard Wood <bwood@gwu.edu> Homo Habilis - Handy Man top
Excavations in 1997 in West Turkana in Kenya have uncovered evidence of stone flake production as early as 2.34 million years ago, several hundred thousand years earlier than previous finds. No hominid remains have been associated with the production sites. If the tool makers were Homo habilis, the dates for this species must be pushed back three hundred thousand years. It is also possible that the tool makers were australopithecines. See H. Roche, A Delagnes, J.-P. Brugal, C. Feibel, M. Kibunjia, V. Mourre, and P.-J. Texier. Early hominid stone tool production and technical skill 2.34 Myr ago in West Turkana, Kenya. Nature 399 (1999): 57-60 (BBC report and abstract). Paleoanthropologists once considered tool-making to be one of the defining characteristics of the genus Homo. Now, the very classification of Homo habilis as part of the genus Homo is questioned; the remains may more appropriately be grouped with the australopithecines. The diversity of tool-making and tool-using behaviors among chimpanzees has forced a complete revision of assumptions surrounding the concept of "man the toolmaker", including revision of ideas concerning the gender of the first tool users. Chimpanzees have diverse and regionally varied repertoires of tool-using, and other "cultural" behaviors. Tool use can no longer be used as a genus criterion. Nor is tool use clear evidence of a more abstract conceptual understanding. Cebus monkeys are considered prolific tool users but exhibit no apparent understanding of cause and effect, or of the difference between appropriate and inappropriate tools. Nevertheless, complex innovations in composite tools remains a very recent phenomenon. With the appearance of near-modern brain size, anatomy, and perhaps of grammatical language approximately 300,000 years ago, the pace of the evolution of human technology quickened exponentially. A mere 12,000 years separate the first bow and arrow from the International Space Station. For a discussion, see S.H. Ambrose's comments in Science 2 Mar 01 291:1748. Migration of homo habilis? top Eitan Tchernov, a palaeontologist at the Hebrew University of
Jerusalem, reports in September '99 that excavations at the
Erq-Lel-Ahmar site in Israel's Jordan Valley have revealed the
remains of thousands of chopping tools, crude axes and stone knives
thought to have been used by prehistoric human beings. They are dated
to two million years -- 500,000 years before the first hominid (homo
erectus) has been thought to leave Africa. The likely candidate
behind the tools is homo habilis, not previously thought to have
engaged in migrations. The results don't appear to have been
published in scientific journals yet; see newspaper
report. Homo Ergaster top Homo ergaster and Homo erectus are the two immediate precursors of Homo sapiens. Homo ergaster is believed to have originated in Africa and to have given rise to H. erectus in Asia. As with other hominid fossil groups, precise evolutionary sequences and geographical loci continue to be debated. For recent finds in Georgia -- the earliest outside of Africa -- dating back 1.7 million years, see BBC news report (11 May 2000). Homo Erectus - Java and Peking Man top First discovered by Eugene Dubois in 1891 in Indonesia, this
fossil group is currently viewed as the closest precursor to H.
sapiens. It was initially given the names "Anthropithecus erectus"
and "Pithecanthropus erectus" and seen to straddle the line between
ape and human. Dubois' species and subsequent finds in China from
1927 ("Peking man") were in 1951 subsumed under the single category
Homo erectus. It is now recognized as a widespread species exhibiting
significant geographical variation (for a discussion, see Jeffrey H. Schwartz in Science 2004 305:53 -- excerpts):
Most remains suggest a stocky build. However, the Turkana Boy
(KNM-WT-15000) is estimated to be a youngster who would have grown to
be about 6 ft tall; he is fairly slender. ER 1808 is another tall
erectus, thought to be about 5'9" (175 cm), and possibly female. See
Walker and Leakey (1993).
Early Homo Erectus Stone Tools in China top Stone tools dated to 1.36 million years ago provide the earliest evidence yet of human occupation of northeast Asia. Full news report (BBC 26 September 2001, external) Did Homo Erectus Use Watercraft 800,000 Years Ago? top Morwood et al. (1998) examined stone tools from two fossil sites on the Wallacean island of Flores, between Southeast Asia and Australia, dating them to 800,000 to 900,000. (They used fission-track dating in zircon grains - a method of dating geological specimens by counting the radiation-damage tracks produced by spontaneous fission of uranium impurities.) Flores has been an island for a least a million years, so the toolmakers - Homo erectus - must have used watercraft to reach it. For a recent study that reaches similar conclusions, see O'Sullivan et al. Geology 29, 607 (2001). The rafting may have been involuntary; as paleontologist Tim White has pointed out, people were rescued from large floating rafts of natural debris after the 2004 tsunami in the area. There is some debate about where this human ancestor arose, some favoring Africa and others Asia; cf. the multiregionalism debate below. What is certain is that Homo erectus used fire and stone tools and appeared around 1.7 million years ago. The species disappeared from Africa around half a million years ago, but remained in Asia for another 250,000 years (and possibly much longer). Morwood et al. (1998) suggest this evidence, combined with the geographical radiation of Homo erectus in the Early Pleistocene (2 million to 10,000 BP; cf. Geological Time-scales) and other recent discoveries, indicates the cognitive capabilities of this species may be due for reappraisal. See Gibbons (1998). For more material, see Mark Rose, First Mariners. Archaeology Magazine 51. 3 (May/June 1998) (external). Contact: M.J. Morwood (mmorwood@metz.une.edu.au) Was erectus contemporary with modern homo in Java? top Recent evidence suggests that homo erectus may have survived much longer than the current consensus suggests. Swisher et al. (1996) report that recent dating of erectus remains from Java place them between 27,000 years and 53,000 years ago. This is a million years after erectus is thought to have died out in Africa and raises the possibility erectus coexisted with modern homo sapiens in Southeast Asia. The erectus population on Java would have been cut off from the mainland for long periods of time by rising seas (though see below for the possibility of erectus watercraft). Isolated from the Eurasian gene flow, they would have been able to survive unchanged until modern humans arrived by boat around 40,000 years ago. See also Gibbons (1996). For a discussion of the implications, see Dan Barnes on Recent Dates
for Javanese Homo erectus followed by the AP report with
interviews, and the multiregionalist Milfred Wolpert's
response, all on the online Human Origin News (external).
Note that Wolpert argues the remains are of homo sapiens, not
erectus; this controversy is tied to the multiregional hypothesis (below). Brown et al. (2004) and Morwood et al. (2004) report in Nature (28 October 2004) the discovery of a new species of homo, a variant of homo erectus, that stood one meter tall and coexisted with modern humans for tens of thousands of years. The find was made on the Indonesian island of Flores and dated to 38,000 to 18,000 years ago. The remains include a nearly intact hominin cranium and some post-cranial bones, likely of a 30-year-old female, along with skeltal remains from several individuals. At 380cc, their brain size was about that of a chimpanzee, yet they manufactured fine stone tools. H. floresiensis is proposed to descend from Homo Erectus populations that may have arrived on Flores as early as 900,000 years ago (see above). This would make them the first case of island dwarfing observed in a hominin species, though this is common in other species. The Flores hominins hunted dwarf mammuth (Stegodon), giant rats, and Komodo dragons. They may have survived into modern times. For details, see the Nature Special Report (external). For commentary, see ScienceWeek 2004-12-17 (external); skulls and proposed lineage at Phyaryngula (external). The stone tools associated with the finds include carefully crafted slicers and other tools indistinguishable from those made by modern homo
sapiens, entirely unlike the multipurpose choppers and scrapers characteristic of homo erectus sites. There is evidence of collaborative
large-game hunting. The extremely diminutive brain volume heightens the anomaly. Additional finds reported by Marwood
et al. (2005) aim to rule out the competing explanation that the remains are microcephalic or otherwise pathological. The authors argue the new
species shares tooth size and facial features with the genus Homo, but body proportions with Australopithecus, and conclude the genealogy remains
uncertain. For a discussion, see Dalton (2005) and Lieberman (2005). Martin et al. (2006) argue the tiny cranial capacity cannot result from normal
dwarfing. The presence of archaic features in a recent population makes this a senstational discovery. A study by Jungers and Tocheri (2009) finds that a well-preserved foot shares features with the Australopithecines. This should put to rest the debate over the authenticity of the finds; they clearly represent a new species, and not a few diseased individuals. The morphology, however, is extremely hard to reconcile with the associated toolset. Homo Erectus Dwelling Discovered in Japan top Japanese archaeologists report in the New Scientist (March 2000) that they have uncovered the remains of what is believed to be the world's oldest artificial structure, on a hillside at Chichibu, north of Tokyo. The site has been dated to half a million years ago, suggesting its architect was homo erectus. It consists of what appear to be ten post holes, forming two irregular pentagons which may be the remains of two huts. Thirty stone tools were also found scattered around the site. The shallow post holes were dug in a layer of volcanic ash, which can be reliably dated. The holes were clearly distinct from the volcanic layer, says Kazutaka Shimada, curator of the Meiji University Museum in Tokyo. "They had well-defined edges." Before this discovery, the oldest remains of a structure were those at Terra Amata in France, from around 200,000 to 400,000 years ago; see artist's conception. Literature on Homo Erectus Brown, F., Harris, J., Leakey,
R. & Walker, A. (1985). Early Homo erectus skeleton from west
Lake Turkana, Kenya. Nature 316: 788-792. Brown, P., T. Sutikna,
M.J. Morwood, R.P. Soejono, Jatmiko, E. Wayhu Saptomo, and Rokus Awe
Due (2004). A new small-bodied hominin from the Late Pleistocene of
Flores, Indonesia. Nature 431:1055-1061. Gibbons, Ann (1996). Human origins - Homo erectus in Java - a 250,000-year anachronism. Editorial. Science 274, 5294 (Dec 13): 1841-1842. Gibbons, Ann (1998).
Paleoanthropology - Ancient island tools suggest Homo erectus was a
seafarer. Editorial. Science 279, 5357 (March 13):
1635-1637. Morwood, MJ et al. (2005). Further evidence for small-bodied hominins from the Late Pleistocene of Flores, Indonesia. Nature 437 (13 October 2005): 1012-1017. Morwood, MJ, PB O'Sullivan, F.
Aziz, A. Raza (1998). Fission-track ages of stone tools and fossils
on the east Indonesian island of Flores. Nature 392, 6672
(March 12): 173-176. Morwood, MJ, R.P.
Soejono, R.G. Roberts, T. Sutikna, C.S.M. Turney, K.E. Westaway, W.J.
Rink, J.-X. Zhao, G.D. van den Bergh, Rokus
Awe Due, D.R. Hobbs, M.W. Moore, M.I. Bird, and L.K. Fifield (2004).
Archaeology and age of a new hominin from Flores in eastern Indonesia. Nature 431: 1087-1091. Swisher III, CC, WJ Rink, SC Antón, HP Schwarcz, GH Curtis, A Suprijo, and Widiasmoro (1996). Latest Homo erectus of Java: Potential contemporaneity with Homo sapiens in Southeast Asia. Science. 274 (5294): 1870-4. Walker, A.and Leakey,
R.(1993). The Nariokotome Homo erectus Skeleton.
Springer-Verlag. From Archaic to Modern Homo Sapies top Archaeological
digs Excavations at Cuxton in Kent have unearthed large flint handaxes and a cleaver, dating from around 250,000 years ago; cf. BBC News article (20 June 2006). A well-preserved site in Norfolk, England, is currently being
excavated; cf. BBC
News article (25 June 2002); cf. BBC update (1 Oct 2006). Ongoing excavations at Blombos Cave
(external) in South Africa have uncovered remains from around 90,000
years ago. The bones are anatomically modern. The findings
demonstrate a rich cultural life and early technological innovations.
For a journalistic take, see Stone Age man
wasn't so dumb (Feb. 2000). Excavations in Skhul Cave on the slopes of Mount Carmel in
Israel and the site of Oued Djebbana in Algeria have uncovered
perforated snail shells likely used in necklaces, dating from
90-100kya; cf. BBC News article (22 June 2006). Paviland Cave Research Project (1996-1998). Paviland Cave (external), on the south coast of the Gower peninsula, South Wales, United Kingdom, is an Early Upper Palaeolithic (Early Stone Age) archaeological site, dating to roughly 30,000 - 20,000 years ago. It is the richest site of its kind in Britain, with four and a half thousand finds, including worked bone and stone (lithic) tools. Paleolithic Human Population Expansion In Africa top Human populations have undergone dramatic expansions in size, but other than the growth associated with agriculture, the dates and magnitudes of those expansions have never been resolved. Genetic approaches to the study of human population expansions have focused on variation at a single genetic locus, the "control region" of mitochondrial DNA. But in the study of demographic history, single-locus investigations suffer from pronounced statistical and biological limitations. The statistical problem is that the conclusions rely on only one particular realization of a gene genealogy, the "tree" determining the ancestral relationships among a set of alleles. The biological problem is that there are a large number of functional genes in the mitochondrion, and due to a complete linkage, a selective sweep for any one of the genes may lead to a spurious signal of expansion. Reich and Goldstein (University of Oxford, UK) present two new statistical tests for population expansion, using variation at a number of unlinked genetic markers to study the demographic histories of natural populations. The authors report that analysis of genetic variation in various aboriginal populations throughout the world reveals highly significant evidence for a major human population expansion in Africa, but no evidence of expansion outside of Africa. The inferred African expansion is estimated to have occurred between 49,000 and 640,000 years ago, certainly before the Neolithic expansions, and probably before the splitting of African and non-African populations. The authors suggest that in showing a significant difference between African and non-African populations, their analysis supports the unique role of Africa in human evolutionary history. The authors also suggest that the missing signal in non-African populations may be the result of a population bottleneck associated with the emergence of these populations from Africa, as postulated in the "Out of Africa" model of modern human origins (see below). (Procedings of the National Academy of Sciences, US 95 (7 Jul 98): 8119.) Contact: David B. Goldstein. Woodruff and Gagneaux translated the variation in the DNA sequences within humans, chimpanzees, and gorillas into a measure of genetic diversity. Chimps and gorillas, they found, have roughly equal level of diversity. Humans, however, although we are spread out over a vastly larger geographic area, have a significantly lower level of genetic diversity. Woodruff sums it up succinctly: "There's more diversity in one social group of 55 chimps than in the entire human population." This lack of genetic diversity suggests the human lineage has gone through a population bottleneck at some point in its past history. When this happened or for what reason has not been determined. A plausible earliest date can be set by considering the large differences found between Neanderthal DNA and modern genes (Krings 1997, 1999, and Goodwin 2000). Unless these differences result from a Neanderthal bottleneck, the common ancestor of Neanderthals and modern humans -- Homo erectus -- was genetically far more diverse than current human populations. Since the human-Neanderthal split, according to the genetic analyses so far undertaken, took place around 500,000 and 800,000 years ago, the bottleneck on the human population must have occurred since this split. See Brooks, Martin (1999). Apocalypse then. Our genes show that early humans teetered on the brink of extinction. New Scientist 163. 2199 (14 August): 32. For a colorful overview of the possible effects of climatic and catastrophic events on human evolution, see The Signposts Perspectives: 59,999,999 BC- 51,000 BC. (external). Stanley Ambrose suggests in a recent article in the Journal of Human Evolution that the eruption of Mount Toba in Sumatra around 71,000 years ago may have caused a worldwide bottleneck in human populations. See abstract. In response to Martin Brooks' "Apocalypse then," Mike Weale at the Centre for Genetic Anthropology, University College London, argues in a recent letter (external) to the New Scientist (4 September 1999) that "there are plenty of other ways to explain why the diversity of human mitochondrial DNA is low compared to that of chimps and gorillas": A recent selective sweep is one answer, in which a new DNA molecule with some selective advantage replaced all others in the population. Another is simply chance. Studies on other parts of the genome, such as the Y chromosome, will help to resolve this issue. Even if a common population bottleneck is indicated for all genes, this need not mean these were the only humans alive at the time. All it means is that any other humans were replaced without contributing to the gene pool when our ancestors, possessing perhaps some cultural innovation, multiplied. Genocide? Possibly, although competitive exclusion is an alternative explanation. Either way, the low diversity of our mitochondrial DNA may indicate too much success rather than near failure.Peter Frost notes that a recent study by Cargill et al. (1999) shows that genetic variability is inversely related to the gene's selective value. Variability is highest at "degenerate" sites, lower in non-coding regions, and lowest in coding regions. So the lower the selective value of a gene, the more it will accumulate "junk" variability. The higher its selective value, the more this "junk" variability will be eliminated. Somewhat counterintuitively, a low degree of variation in the human population could be the result of several regional mutations that confer a selective advantage, spreading through the whole population in a manner Brace calls mosaic evolution. Instead of tidy lineages and population replacements, we get a situation where gene flow in effect creates a single (if still varied) but widely dispersed population. Cargill, M. et al. 1999. "Characterization of single-nucleotide
polymorphisms in coding regions of human genes." Nature
Genetics 22: 231-238. The earliest indirect evidence of clothing is argued by
Dr.
Mark Stoneking, of the
Institute for Evolutionary Anthropology in Leipzig, to be traceable in
the genetics of the human body louse. This louse, which lives only in
clothing, not hair, split from the human head louse some time between
72,000 and 42,000 years
ago. (Modern humans are thought to have entered Europe around
40,000 years ago.) It's unclear if the body louse lives in furs or only
in textiles. Early Evidence of Art Archaeologists in Zambia have uncovered evidence that early humans
used paint for aesthetic purposes far earlier than previously thought
(see BBC
report, 2 May 2000). The team found pigments and paint grinding
equipment dated to 350 - 400 kya; the previously oldest pigments are
120,000 years old. Over 300 fragments of pigment have now been
uncovered in a cave at Twin Rivers, near Lusaka, Zambia, originally
gathered from the surrounding area. One of the team who made the
discovery, Dr Lawrence Barham from the University of Bristol, said:
"We're dealing here with people who were perhaps using symbols far
earlier than we expected." He speculates "It also implies the use of
language, so it's an important discovery, full of implications for
the development of new behaviours." Composite tools of more than one
type of material -- for instance wood and stone -- also date from
this period; cf. Mithen's (1996) notion
of cognitive fluidity. Homo Sapiens Idaltu The origin of anatomically modern Homo sapiens and the fate of Neanderthals have been fundamental questions in human evolutionary studies for over a century. A key barrier to the resolution of these questions has been the lack of substantial and accurately dated African hominid fossils from between 100,000 and 300,000 years ago. Here we describe fossilized hominid crania from Herto, Middle Awash, Ethiopia, that fill this gap and provide crucial evidence on the location, timing and contextual circumstances of the emergence of Homo sapiens. Radioisotopically dated to between 160,000 and 154,000 years ago, these new fossils predate classic Neanderthals and lack their derived features. The Herto hominids are morphologically and chronologically intermediate between archaic African fossils and later anatomically modern Late Pleistocene humans. They therefore represent the probable immediate ancestors of anatomically modern humans. Their anatomy and antiquity constitute strong evidence of modern-human emergence in Africa. The fossils, which are assigned to a new subspecies, Homo sapiens idaltu, "predate classic Neanderthals and lack their derived
features" (White 2003), suggesting modern humans evolved independently of the Neanderthals. The best preserved of the three Idaltu crania has a
spacious braincase, with a cranial capacity around 1,450cc, at the high end of the modern range. Several features, such as relatively heavy brow
ridges, "greater craniofacial robusticity, greater anterior–posterior cranial length, and large glenoid-to-occlusal plane distance" (White
2003), deviate from modern measurements, but represent a clearly different lineage than the Neanderthals. See one view of the holotype skull of the new paleosubspecies and artist's reconstruction. The remains were embedded in archaeological assemblages containing "elements of both Acheulean and Middle Stone Age technocomplexes.
Associated faunal remains indicate repeated, systematic butchery of hippopotamus carcasses." (Clark 2003). Clark, J. D. et al. (2003). Stratigraphic, chronological and behavioural contexts of Pleistocene Homo sapiens from Middle Awash, Ethiopia. Nature 423: 747-752. Neanderthals top About 10 kilometers east of Dusseldorf in Germany, in the valley
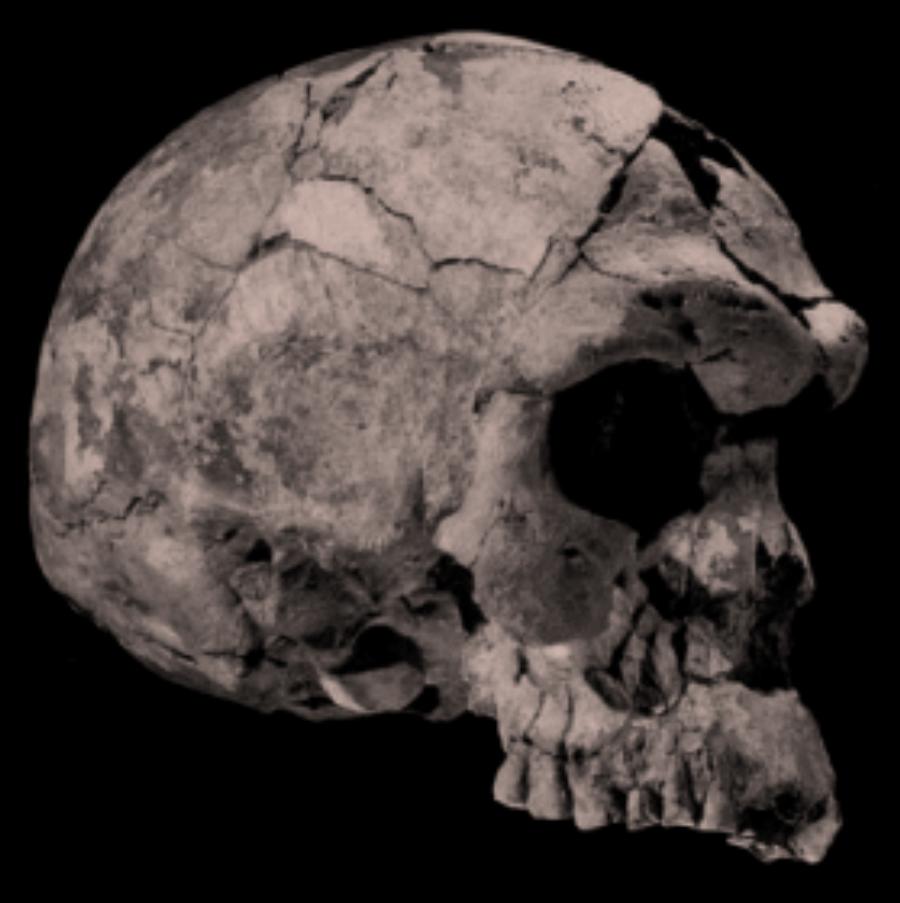
of the Dussel, there is a small town called Neander. One hundred and
forty-three years ago, in the summer of 1856, some workmen broke into
a cave to get at the limestone inside and discovered a set of ancient
bones. Most of the bones were smashed to bits by the workmen, but
some of the bones, including part of the skull, survived, and the
skeleton was soon recognized by anthropologists as belonging to an
ancient race of men who came to be known as the Neanderthals. A
Neanderthal fossil had actually been discovered some years earlier in
Gibraltar, but not recognized as such. Neanderthal-like fossils have
also been found in France, Spain, Italy, Yugoslavia, Iraq, and
Israel.
The shaded area indicates the known range of Neanderthals. Finds of archaic Homo in China and Java are also classified by some researchers as Neanderthal. Mezmaiskaya is the location where the baby Neanderthal whose DNA was recently sequenced was found (see Goodwin et al. 2000); the earlier Neanderthal DNA sequence examined by Krings et al. (1999) was determined from bones found in Feldhofer Cave in Germany (see below).For more than a century, one of the central questions in paleoanthropology has been whether modern man evolved from this hominid -- or was the Neanderthal a separate branch that became extinct? In general, "Neanderthals" is the informal designation of a morphologically distinctive group of large-brained hominids who inhabited Europe and western Asia between approximately 200,000 and less than 30,000 years ago. They are sharply distinguished from modern humans by a wide range of cranial and postcranial characteristics, although they do share a number of derived bony features with other members of the European/western Asian hominid clade that diversified in this part of the world after approximately 500,000 years ago. Subsequent to approximately 150,000 years ago, the Neanderthals appear to have been the sole surviving species of this clade. Some researchers describe them as a robust species of homo occupying cold climates in Europe and Western Asia from around 200 to 30 ka bp. Their brain volume was slightly larger than the contemporary average - cf. Neanderthal skulls. The Neanderthals were apparently highly successful over a large
region for a substantial period of time, but this situation changed
dramatically with the arrival in Europe of the first modern humans,
Homo sapiens. The evidence is that these "Cro-Magnons" had begun to
arrive both in eastern Europe and in the far northeast of the Iberian
Peninsula by approximately 40,000 years ago, and within little more
than 10,000 years, the Neanderthals were gone (for recent finds in
central Europe dating to 28,000 kya, see "The
Latest Neanderthal" (SciAm, external)). The mechanism of
their eviction has long been debated, but there are four main
possibilities. The first and second of these possibilities, that the
Neanderthals were eliminated by the moderns in direct conflict (cf.
a proposed
scenario and map in Der Spiegel, March 2000; external, and
Valerius Geist's response) or
by indirect economic competition, both imply the separate species
status of the Neanderthals, as does any combination of these two
possibilities. The alternative possibilities, that the Neanderthals
had simply evolved rapidly into moderns, or that the genes of the
invading moderns simply "swamped" those of the Neanderthals, both
imply some form of species continuity (cf. Loring Brace, below). Francisco J. Jiménez-Espejo and colleagues (2007) have argued a brief spell of sharply cooler climate is a more promising candidate for the final Neanderthal die-off than competition with modern humans; see also arguments byClive Finlayson to this effect. In a recent article, Simon Y. W. Ho and colleagues (2005)
argue that analyses of molecular rates of genetic change have led to
systematically overestimated divergence times. In brief, they
suggest it is not useful to apply a single short-term or long-term
rate, but that the data must be interpreted according to a rate decay
curve to reflect the transition from a high short-term rate to a low
long-term rate. Their recalibration suggests that the ancestors
of modern humans and Neanderthals diverged around 354 kya
(222-705 kya), and that the last common ancestor of all Neanderthal
sequences lived around 108,000 years ago (70-156 kya). They also
arrive at a far more recent date for the last common ancestor of all
humans at 76,000 years ago (47–110 kya). In 2006, Richard E. Green et al. examined a 38,000-year-old
Neanderthal fossil that is exceptionally free of contamination from
modern human DNA. They write, "sequencing of a DNA extract from this
fossil has thus far yielded over one million base pairs of hominoid
nuclear DNA sequences. Comparison with the human and chimpanzee genomes
reveals that modern human and Neanderthal DNA sequences diverged on
average about 500,000 years ago." For details, see the Nature web focus on Neanderthal DNA. Richard E. Green et al. (2006). Analysis of one million base pairs of Neanderthal DNA. Nature 444, 330-336 (16 November 2006). Neanderthals and the Evolution of
Language top A topic of much debate is the date for the evolution of language. In part, there is a definitional problem: while vervets use vocal cries that share many of the characteristics of words, and bonobos (more so than chimpanzees) can be trained to communicate with signs. The female bonobo Kanzi, for instance, one of the primary subjects in E. Sue Savage-Rumbaugh's research, has demonstrated impressive symbolic abilities. In her SRCD Monograph, Savage-Rumbaugh has documented that Kanzi could comprehend more than 440 novel sentences of request and performed equal to or somewhat better than did a 2-1/2 yr old girl tested in the same manner and with the same sentences. Kanzi's language production was assessed by Patricia Greenfield of UCLA at the level of a 1-1/2 yr old child. (Contact: Sue Savage-Rumbaugh and Duane Rumbaugh <LRCDMR@langate.gsu.edu>.) This of course still leaves a considerable distance between the linguistic capabilities and performance of human beings and that of our closest relatives. However, it is not in principle difficult to imagine forms of speech intermediate to ours in complexity; for instance, speech with a reduced set of phonemes, grammatical categories, and sentence forms. The difficulty is determining what kinds of evidence are relevant to this question, and how to evaluate the scarce clues available to us. Cultural evidence top It is no doubt the case that achaeological remains of culture can tell us something about the cognitive abilities of the people in question. However, in order to interpret the cultural remains, we need a theory of how various cognitive abilities enable (and are required to account for) certain cultural activities. At present, there is no clear consensus on such a theory. In the context of language, we specifically need a model of how the biological and cultural innovations of language - and they must of necessity go hand in hand - impact our cognitive skills. Is it for instance the case, as Merlin Donald (1991) argues, that human beings today can function very well without language, retaining their abilities to interpret events, interact socially, plan their days, and communicate through a range of mimetic means? Or, on the contrary, are human minds generated by language in a radical way, so that deprived of language, we descend to the cognitive skills of chimpanzees? This view might be suggested by the apparently severely reduced cognitive capacities of children raised by animals. It is also a view that resonates with the idea that human beings are caught in the "prison-house of language" (Nietzsche), depending for their basic cognitive skills on the culturally constructed nature of language. In my view, it is fruitful to examine the cognitive abilities
associated with the imagination as more fundamental to advances in
human cognition than language per se. Such advances are required for
the emergence of certain forms of language, notably those that do not
refer directly to what is present. Thus, evidence of imagination can
be taken as a minimal requirement for language as internal thought.
However, the converse does not necessarily hold: internal thought
does not appear to require language. The flower-bedecked grave at Shanidar top Earlier claims of a Neanderthal flower burial in Shanidar is now
contested; see Gargett, R.H. (1989) Grave shortcomings: The evidence
for Neanderthal burials. Current Anthropology. 30. 157-90. The
precise implications of such a find also need to be argued and
spelled out in detail. The hyoid bone top Evidence that Neanderthals had a morphologically near-modern vocal
apparatus is presented in Arensburg, B., Tillier, A.M.,
Vandermeersch, B., Duday, H., Schepartz, L.A. and Rak, Y. (1989). A
middle palaeolithic human hyoid bone. Nature 338: 758-760. It
is depicted to scale in Donald C. Johanson and B. Edgar's From
Lucy to Language (Simon and Schuster, 1996), p. 107. For a
discussion, see Arensburg, B., Schepartz, L.A., Tillier, A.-M.,
Vandermeersch, B., Duday, H. & The hypoglossal canal top Kay, Cartmill, and Balow at Duke have recently measured the relative size of the hypoglossal canal in primates. This canal, a hole at the base of the skull in the area where the spinal cord enters the head, is the conduit for the nerve fibers from the brain to the muscles of the tongue, and an interesting index of complex speech. See the Duke University press release (external) and John Wilford's presentation of the results in The New York Times, April 28, 1998. In brief, the findings indicate that the enlargement of the hypoglossal canal to modern proportions happened already in early forms of archaic homo sapiens around 400,000 years ago. Neanderthals show the same-sized canal and according to this line of evidence would have possessed complex speech. Because of their shorter neck, their range of vowels may have been narrower. More recently, this line of reasoning has been challenged; cross-species comparisons suggest the hypoglossal canal may not be a meaningful indicator of adaptations for speech. See Kay et al. (1998). Contact: Matt Cartmill, Biological Anthropology and Anatomy, Duke University, NC. For evidence that the Neanderthal were poor at cultural
innovation, yet capable of acquiring novel technology from their
Homo sapiens sapiens neighbors, see Paul Mellars, The
Neanderthal legacy: an archaeological perspective from western
Europe (Princeton, NJ: Princeton University Press, 1996). Literature on Archaic Homo top For a recent review, see Strait, DS; Grine, FE; Moniz, MA. (1997). A reappraisal of early hominid phylogeny. Journal of Human Evolution. 32 (1): 17-82. deCastro, JMB; Arsuaga, JL; Carbonell, E; Rosas, A; and others (1997). A hominid from the lower Pleistocene of Atapuerca, Spain: Possible ancestor to Neanderthals and modern humans. Science 276 (5317):1392-1395. Gabunia, L. & Vekua, A. (1995) A Plio-Pleistocene hominid from Dmanisi, East Georgia, Caucasus. Nature. 373: 509-12. Grün, R., Brink, J.S., Spooner, N.A., Taylor, L., Stringer, C.B., Fransiscus, R.G. & Murray, A.S. (1996) Direct dating of Florisbad hominid. Nature. 382 (6591): 500-1. McDermott, F., Stringer, C.B., Grün, R., Williams, C.T., Din, V.K. & Hawkesworth, C.J. (1996). New Late-Pleistocene Uranium-Thorium and ESR dates for the Singa hominid (Sudan). Journal of Human Evolution 31 (6): 507-516. Mellars, Paul. Origins of Anatomically Modern Humans. Journal of Field Archaeology 24, 3 (Fall, 1997):377 (4 pages - book reviews). Rightmire, G.P. (1996) The human cranium from Bodo, Ethiopia:
Evidence for speciation in the Middle Pleistocene. Journal of
Human Evolution. 31: 21-39. Ronen, A. and Weinstein-Evron, M. (2000). Toward modern humans: the Yabrudian and Micoquian, 400-50 k-years ago. Proceedings of a congress held at the University of Haifa, November 3-9, 1996. Oxford: Archaeopress, 2000. Tautavel Man. A web presentation by the French Ministry of Culture of an archaic homo found in Southern France in 1971 and dated to 450,000 years. Full text and illustrations (external). Turner, A. (1992) Large carnivores and earliest European hominds:
Changing determinants of resource availability during the Lower and
Middle Pleistocene. Journal of Human Evolution. 22 (2):
109-26. Out of Africa versus
Multiregionalism top
Archaeological finds from Java to Kenya demonstrates with certainty that Homo erectus inhabited a vast territory extending through Eurasia and Africa. Appearing in the lower pleistocene, around two million years ago (cf. Geological Time-scales), the remains are initially confined to the tropics, but Homo erectus soon spread into temperate zones. Its toolkit was simple but adequate for this first of hominids to join the league of major predators. Somewhere around 250 000 years ago, we begin to see changes in
skull sizes in Africa. Over the next 100,000 years, there are
numerous finds of so-called Archaic Homo sapiens (see literature above). Tim White's team announced in
June 2003 a series of finds in Ethopia that look modern; they are
confidently dated to around 160,000 years ago (see Homo sapiens idaltu).
The first Levallois flint tools appear in southern Africa around
120,000 years ago. In the Levant, various populations with a broad
range of modern and archaic features coexist in the period between
120,000 and 60,000. Although early humans ventured out of Africa
briefly, as seen in the 90,000-year-old fossils found at Skhul and
Qafzeh in Israel,they do not appear to have gained a permanent foothold in the Middle East or Europe until much later.
The earliest human remains outside of Africa and the Levant are at Lake Mungo in Australia; early estimates of dates older than 60,000 years ago have been adjusted to around 46,000 (for references, see Forster & Matsumura, 2005). The Niah Cave of
Borneo was inhabited around the same time, >45,000 years ago. A new analysis of mtDNA from Malaysians and
aboriginal Andaman islanders (Forster & Matsumura, 2005)
indicate that a small number of hunter-gatherers left Africa around
65,000 years ago, following the coastline of the Indian Ocean, and that
their descendants fanned out into the rest of the world.
Archaeological and genetic data suggests fully modern peoples did not
move into Europe until some 40,000 years ago (Mellars 1992), possibly because it was
already inhabited by Neanderthals. For more details on the early migrations, see below. While recent genetic evidence converges on an African origin of this latest wave of fully modern human migrations, it has long been debated where the earliest East-Asian populations originated. Did Homo sapiens emerge in a single speciation event in Africa some time around 150,000 years ago, expanding into the territory of Homo erectus and replacing it? Or did local populations of Homo erectus, spread through the breadth of the vast Eurasian and African continents, evolve together into Homo sapiens, mixing genes and also retaining local characteristics? Archaeologists working in the field for a long time leaned towards the latter scenario, suggesting that human evolution shows regional continuity (the so-called Multiregional model). Europe and Western Asia posed a special case, as it was populated by Neanderthals, a form of archaic Homo that by some researcherswas thought to be the ancestor of the subsequent modern human population (see above for background and below for discussion). During the late 1980s, initial studies of global DNA diversity at a stroke changed this picture. The much greater amount of genetic variation in Africa pointed clearly to that continent as the locus of the first fully modern human populations. In a landmark paper, Cahn, Stoneking, and Wilson presented an analysis of a sampling of human mitochondrial DNA — inherited only through the maternal line — from around the globe. They argued that the roots from the "most parsimonious tree" that described the phylogenetic relationship of human mitochondria had deep roots in Africa, the next deepest roots were in Asia, followed by European and New World lineages. Using a mitochondrial clock, they computed the time at which all these mitchondria coalesced into a common ancestor — the Mitochondrial Eve. She lived, they argued, some 180,000-270,000 years ago in North Africa. In this "out-of-Africa" scenario, modern humans arose in Africa
around 150,000 years ago and then spread elsewhere, replacing
Neandertals and Homo erectus -- and any other hominid relatives in
our evolutionary past. A Japanese team estimated that all
non-Africans are descendants of a small band of humans that left
Africa around 100,000 years ago; Forster & Matsumura (2005) have now moved this date forward to around 65,000 years ago. In May 2001, R. Spencer Wells et al.
reported that their examination of the Y-chromosome in Neanderthals shows no trace of
any non-African material: "This really puts the nail in the coffin of
multiregionalism" (BBC
report, external). Decades of anthropological debate over modern
human origins based on interpretations of measurements of limb
proportions and a host of bumps and grooves on ancient fossils
teetered on the verge of irrelevance. Continuity in South-east Asia? top There are, however, difficulties with the model. Alan Templeton took the data from Cahn, Stoneking and Wilson's paper and showed that the number of trees that could be constructed from their data was astronomical. Their "most parsimonious tree" was not unique; in fact, Templeton found even more parsimonious trees with strikingly different topologies. These suggested the possibility of a continuous and distributed origin of modern homo sapiens, fueled by a continuous exchange of genes. This lead to the construction of the Multi-regional hypothesis for the origins of modern homo. In this view, the evolution and diversification of modern homo sapiens took place across a large geographic region that encompassed Africa, Asia, and Europe. Excehange of genes through migration took place during the 200,000+ years of modern homonid evolution. There is no single cradle of origin but a "thousand points of light." The hypothesis appeared to find the strongest support in South-East Asia. Paleoanthropologist Loring Brace (1995) points out that the out-of-Africa hypothesis is not supported by much of the archaeological evidence. Most strikingly, there is no detectable break in cultural development in South-east Asia that suggests the replacement of Homo erectus by migrating Homo sapiens. The mtDNA analysis relies on a steady trickle of mutations -- the so-called genetic clock; this assumption may be flawed. Further genetic data could also turn the tide: recent examinations of genetic variation in New Guinea -- for a preview, see Redd & Stoneking (1998) -- finds that it is comparable to that of Sub-Saharan Africa; this would seem to indicate a longer history than that presumed by the out-of-Africa hypothesis, and could be explained by a continuity with local Homo erectus populations. In addition, data obtained by Mike Hammer and colleagues on Y-chromosome variation ties New Guinea and Australia to Indonesia and eastern Asia with a date that goes far enough back into the Pleistocene that Homo erectus becomes the predicted progenitor (cf. Hammer 1996 and Karafet 1997; as of July 1999, not all the relevant data has been published). Brace's extensive craniofacial data support this view; according to him, there are similarities in skull features between Asian Homo erectus populations and modern Asians. The discovery (May 1999) that the gracile Australian Mungo Man is as old as 62,000 years has added to the complexity of this debate; this population may have merged with the more robust peoples arriving later (see Peter Brown's Australian paleoanthropology index (external). Thorne's DNA analysis of Mungo Man (January 2001) supports this scenario. The results indicate that the gracile population that first settled Australia 60,000 or earlier are not descendants of a recent wave out of Africa. They also indicate that significant genetic variation continued to exist in the region; around 20,000 years ago, Australia may have seen the arrival of a robust-looking population of homo, which subsequently mixed and merged with the local gracile-looking population.
According to Milford Wolpoff (1997), the fundamental claim of multiregional evolution is that excepting extinctions, ancient populations have multiple descendants and modern populations have multiple ancestors. In Brace's view, novel adaptations can spread efficiently across the Old World without the need for much population movement. Given even the tiniest trickle of a gene flow, natural selection will ensure that novel genes that have an adaptive advantage spread through the population. This process would leave the rest of the phenotype unchanged, thus preserving a large number of regional characteristics -- what Brace calls mosaic evolution. Instead of replacement, we get evolutionary continuity in each region, leading to different races. These races do not differ in significant functional adaptations, but show variation in matters that have little significance for survival, such as the specific proportions of the face, the texture of the hair, or the color of the skin. For the application of genetics to the question of race, see K. Owens and M-C. King (1999), Genomic views of human history. If evolution is continuous and there is no replacement, why are
modern human crania distinct from their generally more robust archaic
predecessors? Brace argues that steady reductions in the size of
canines over the past 100,000 years may be due to mutations in the
parts of the genes that produce the relevant growth enzymes; small
changes here would make the enzyme less efficient and lead to smaller
teeth. As long as the larger teeth are not selected for by
evolutionary pressures, there will be a slow drift towards smaller
teeth. Thus, it is cooking innovations that are responsible for the
reduction of tooth size. Around ten thousand years ago, the selective
forces on teeth drops dramatically with the invention of pottery and
the possibility of surviving on liquid food; before this period,
there are no instances of toothless individuals, but they become
common subsequent to this date. One of Brace's former students,
Margaret J. Schoeninger, wrote her dissertation on changing amounts
of strontium in Archaic and Modern Homo Sapiens as an index of meat
eating: while an archaic diet consisted of 40-60% meat, this fell
dramatically around 18,000 years ago in the Middle East. Primitive
sickles from the period confirm an increased reliance upon grains,
although this is eight to ten thousand years before agriculture (Schoeninger 1980). Challenging the Replacement Hypothesis in
Europe top The replacement hypothesis has also been applied to Europe, where
people from Asia and the Middle East are cast as newcomers into Neanderthal
territory, displacing them completely over a period of some five
thousand years (a view held by Cambridge archaeologist Paul
Mellars and a majority of other researchers in the field). If we look at the European mtDNA gene pool, we find that no
lineages go back any further than 50,000 years (Richards et al 1996); this time depth
corresponds roughly to the entry of the Aurignacian culture into
Europe from the Middle East (Mellars
1992). Recent comparative analyses of contemporary human and
fossil Neanderthal DNA indicates that Neanderthals split from our
ancestors as early as 700,000 years ago, and that there was no
subsequent interbreeding (Krings 1997 and 1999). Again, the analysis was done
on a small part of mDNA, and relies on a genetic clock argument.
Loring Brace argues instead that Neanderthals simply evolved into modern northern Europeans. His skull measurements show that Neanderthals cluster with one and only one modern population: that of Britain and Scandinavia. Robust features of the skeleton, such as brow ridges, simply vanish when the selective force of evolution is no longer active. That the process of that reduction in robustness by which modern appearance arose, he argues, is why virtually all early Upper Paleolithic groups present a kind of 'mixed' appearance. This story can be told with great precision using data from teeth. Brace writes, "As soon as earth oven cookery became a standard part of Neanderthal food-preparation practices coincident with the onset of the last glaciation, tooth size began to reduce at the rate of one percent every two-thousand years. There was a reduction of about 30% of total tooth size between the early Neanderthals of around 130,000 years ago and the "classic" Neanderthals of 50 to 70,000 years ago. Then between 50 and 35,000 years, there was another 10% drop to the point where there is virtually no difference in size between the last people being called Neanderthals (and really more because of stone tool typology than what they actually looked like) and the oldest Upper Paleolithic specimens. And if one looks at the shoulder reinforcements of those same specimens, they are perfectly intermediate between earlier full Neanderthal form and that of living Europeans. Tooth size continued to drop and the difference in tooth size in living Europeans and the early Upper Paleolithic is greater than the difference between the latter and the classic Neanderthals. The assumptions of two distinct populations living in different areas is a product of mind set and not of the data." These dramatically different models -- and the genetic evidence still favors the Out of Africa hypothesis -- both require further data before the debate is over. A more extensive analysis of Neanderthal genes beyond mitochondrial DNA would go some way to clear up the question. Further fossil finds may also throw light on the debate. In April 1999, Portugese archeologists reported the discovery of a hybrid Neanderthal-Cro Magnon child dating from 4,000 years after the commonly agreed date of the disappearance of full Neanderthals (see media report (local) and Discover article (external)). The view that the skeleton represents a hybrid is contested, however; Brace suggests it is simply another example of the gradual reduction in robustness that was taking place across European Neanderthal populations, which in his view did not disappear but change into modern Europeans. Updates on the multiregionalism debate Bower, Bruce (1999). DNA's Evolutionary Dilemma: Genetic
studies collide with the mystery of human evolution. Science News February 6, 1999. pp.
88-90. See also an update on the African Eve hypothesis (8 September '99). Wong (1999). Is Out Of Africa Going Out The Door? Scientific American, July 1999 (external). Evidence of migration routes (new material May 2005) top
The cumulative weight of the evidence is continuing to tilt in favor of the Out of Africa model, and a scenario of human migrations into Eurasia is now beginning to emerge. The genetic analyses reported in R. Thomson et al. (2000) suggest that the spread of Y chromosomes out of Africa is much more recent than previously believed -- on the order of 50,000 years. For a journalistic presentation, see the BBC report (external) on work by Wells et al. backing up this result. Ke et al. (2001) reach a similar conclusion:
Cf. the subsequent discussion (external, subscription required). Some reservations regarding these results is warranted, as the analysis of the geographical distributions of the human genome is far from complete and the statistical analyses is relatively likely to contain flawed assumptions. Alan Templeton (2002), returning to the multiregionalism debate he may be said to have initiated (see above), notes that new information on the human genome is now available, and a reassessment of the multiregionalism debate is possible. He presents a "formal statistical analysis of human haplotype trees for mitochondrial DNA, Y- chromosomal DNA, two X-linked regions and six autosomal regions," concluding that
I had written in late 2001, "The final nail in the coffin of
multiregionalism is not yet in. Evidence of ancient genetic
continuities from local populations may still surface, even though
several studies now point univocally towards a recent African origin
of all contemporary human beings." Templeton's evidence for repeated
migrations out of Africa, in a general context of ubiquitous
interbreeding, presents an elegant synthesis of the
opposing positions in the debate. His analysis suggests that a full replacement
did not take place, yet crucial new mutations took place in Africa
and flowed out of Africa in at least two major expansions. Yet other analyses of the human genome stubbornly point to a single African origin. Evidence presented by Zhivotovsky, Rosenberg,and Feldman in mid-2003, from an analysis of microsatellite genes in 52 populations worldwide, emphasizes the surprisingly low variance in the human genome. They suggest humanity may be descended from as few as 2000 individuals around 70-140,000 years ago. These results do not rule out additional out-of-Africa migration routes, but lend little support to regional continuity. Zhivotovsky, Noah A. Rosenberg, and Marcus W. Feldman (2003). Features of Evolution and Expansion of Modern Humans, Inferred from Genomewide Microsatellite Markers. American Journal of Human Genetics 72:1171-1186. Abstract and full text (external).A followup series of studies of mitochondrial DNA of indigenous peoples of Malaysia and the Andaman islands by Thangaraj et al. (2005), Macauley et al. (2005), and Forster and Matsumura (2005) further suggests that our African ancestors took an initial southern route along the coastline of the Indian Ocean around 65 thousand years ago: 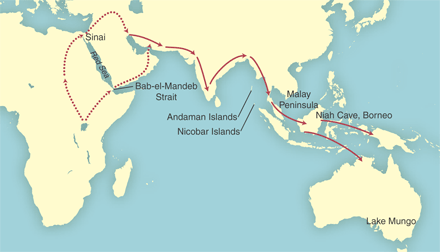 The route may indicate a seafood diet. According to Forster and Matsumura's assessment of the genetic evidence, the rest of Asia, Europe, and the Americas were colonized by descendants of this initial group of migrating early humans. From the mitochondrial data the authors estimate this founding population included only several hundred women. Europe and the Middle East, according to Macauley et al. (2005), was settled from an early offshot of this migration that moved into Europe once the climate improved. Forster, Peter & Matsumura, Shuichi (2005). Did Early Humans Go North or South? Science, Vol 308, Issue 5724, 965-966 , 13 May 2005. Full text (external).This emerging model makes sense of two key features of the current genetic and fossil evidence: a genetic bottleneck in non-African humans dating back to around 70,000 years ago, and the presence of ancient human remains that preserve certain archaic features in South-East Asia.
References on the Multiregionalism debate: Brace, C. Loring (1995). Biocultural interaction and the mechanism of mosaic evolution in the emergence of "modern" morphology. American Anthropologist 97 (4): 711-23. Hammer, Michael F. and Stephen L. Zegura (1996). Role of the Y chromosome in human evolutionary studies. Evolutionary Anthropology 5. 4: 116-134. Karafet, Tatiana, et al. (1997). Y chromosome markers and trans-Bering Strait dispersals. American Journal of Physical Anthropology 102.3: 301-314. Krings, M. et al. (1999). Proc. Nat. Acad. Sci. USA 96: 5581-5585. Krings, M; Stone, A; Schmitz, R W; Krainitzki, H; Stoneking, M; Paabo, S. Neandertal DNA sequences and the origin of modern humans Cell 90 (1997): 19-30. Abstract. Mellars, P.A. (1992). Phil. Trans. Royal Society Lond. B. 337: 225-234. Redd, Alan J. and Mark Stoneking (1998). Evolutionary affinities of aboriginal Australian and Papua New Guinea populations: A comparison of mitochondrial, HLA, and Y chromosome genetic data. American Journal of Physical Anthropology Supplement 26: 185. Richards, M. et al. (1996). Am. J. Hum. Genet. 59: 185-203. Schoeninger, Margaret J. (1980). Changes in Human Subsistence Activities From the Middle Paleolithic to Neolithic Period in the Middle East. U. of Mich. Ph. D. thesis.
Templeton, Alan (2002). Out of Africa again and again. Nature 416 (2002): 45 - 51. Full text (external); excerpt (local). White, Tim, Berhane Asfaw, David Degusta, Henry Gilbert, Gary D. Richards, Gen Suwas, and F. Clark Howell. Pleistocene Homo sapiens from Middle Awash, Ethiopia. Nature 423 (2003): 742-747. Wolpoff, M.H. and R. Caspari (1997). Race and Human
Evolution. Simon and Schuster, New York. Zhivotovsky, Noah A. Rosenberg, and Marcus W. Feldman (2003). Features of Evolution and Expansion of Modern Humans, Inferred from Genomewide Microsatellite Markers. American Journal of Human Genetics 72:1171-1186. Abstract and full text (external). See also news report (BBC, external). |
|
|
|
|---|---|---|
|
|
||
|
|
![]()
© 1998 Francis F. Steen,
Communication Studies, University of California, Los Angeles

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}